Activity
Mon
Wed
Fri
Sun
Aug
Sep
Oct
Nov
Dec
Jan
Feb
Mar
Apr
May
Jun
What is this?
Less
More
Owned by Johannes
Sie suchen in Ihrer Führungsrolle Klarheit? Coaching und Supervision. Systemisch, reflektiert, ressourcenzentriert – und Sie dabei immer souverän.
How to use doubt instead of fighting it – free community to learn Doubt Cybernetics >> usedoubt.com
Memberships
Semantic SEO (Guides & SOPs)
692 members • Free
S K O O L Community Deutsch
675 members • Free
11 contributions to Brain Performance & Competence

Sep '25 •
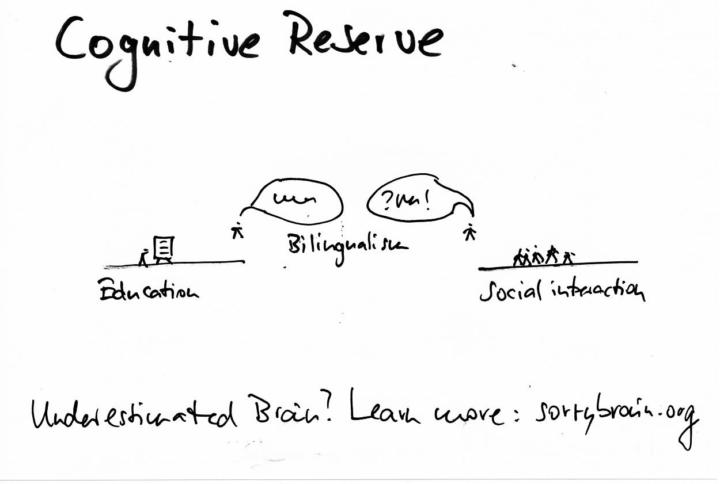
Cognitive reserve from education, social interaction, bilingualism
Cognitive reserve is the capacity to maintain function despite aging or pathology by using more efficient networks, alternative routes and flexible strategies. Education builds representational depth and learning-to-learn skills; social interaction continuously exercises attention, language, memory and emotion regulation; bilingualism trains conflict monitoring and task switching through code-selection demands. Together they strengthen frontoparietal control, language and default-mode couplings, increasing network efficiency at rest and during effort, and supporting compensation when primary circuits falter. Observational cohorts and imaging studies converge on later clinical onset of cognitive impairment with higher reserve, alongside signatures like lower task-evoked effort for the same accuracy, broader recruitment under load and more resilient white-matter pathways. What remains open are causal dose–response curves, the role of socioeconomic confounds and the conditions under which bilingual advantages generalize. Evident in Neuroscience and Brain Research Education functions like repeated strength training for symbolic manipulation and error-guided learning – years of schooling and ongoing study correlate with delayed expression of dementia symptoms, more robust language and executive test scores and leaner hemodynamic responses during problem-solving, a marker of efficient coding. Social interaction is neural cross-training – conversation, turn-taking and perspective-taking recruit hippocampus, temporal-parietal junction, anterior insula and medial prefrontal cortex, while emotionally meaningful ties buffer stress physiology that otherwise harms memory circuits; larger, more active networks predict slower decline and better everyday function. Bilingualism is an in-place executive tutor – selecting one language while inhibiting the other repeatedly engages anterior cingulate and dorsolateral prefrontal cortex, strengthening inhibition, switching and monitoring; lifelong bilinguals often show later clinical onset for similar brain burden, and imaging reveals greater frontoparietal recruitment or thicker left inferior parietal cortex in proficient speakers. Across these levers, reserve looks like three interlocking properties – efficiency (less effort for the same output), capacity (more available circuitry under load) and flexibility (faster reconfiguration when the usual path stumbles). In data terms, think compact EAVs woven into the story – frontoparietal control network – efficiency – higher; language control circuits – flexibility – increased; social network size – attribute – larger predicts steadier cognition – and simple triples your mind can keep: richer education improves network efficiency; active social life supports hippocampal-DMN integration; bilingual practice enhances conflict monitoring and task switching.
1
0
Sep '25 •
Neuroplasticity under trance states: cortical inhibition and associative flexibility
Neuroplasticity under trance states is the coupling of two processes – cortical inhibition and associative flexibility. The first is a protective hush: prefrontal–cingulate control tunes inhibitory interneurons, sensory cortices dampen competing input, and large-scale networks shift toward alpha–theta rhythms that reduce distractor gain. The second is a constructive opening: with noise down, hippocampal–temporal systems and default-mode assemblies can traverse a wider graph of associations, enabling reconsolidation of emotional memories, reframing of self-narratives and creative recombination. Hypnotic analgesia, focused imagery for habit change, and music-induced trance illustrate the same motif – less chatter where it hinders, more linkage where it helps. These states do not magically implant suggestions; they change the signal-to-noise ratio so that targeted learning lands and sticks. Evidence in Neuroscience and Brain Research about cortical inhibition and associative flexibility Across hypnotic and absorption paradigms, imaging repeatedly shows rebalancing among control, salience and default networks. Dorsal anterior cingulate – the brain’s conflict/“error” alarm – often quiets, making intrusive signals less bossy; anterior insula relays interoceptive cues with a calmer precision; dorsolateral and frontopolar prefrontal regions coordinate the task set, keeping the mind inside the guided scene. At the mesoscopic level, alpha and frontal-midline theta increase – signatures of inhibitory gating and top-down stabilization – while occipito-temporal cortices reduce reactivity to irrelevant input. This selective hush shows up behaviorally: under suggestion, pain, itch, or craving cues elicit smaller autonomic spikes, and attention tracks imagined rather than literal stimuli without losing overall coherence. When interference drops, associative travel grows. Hippocampal replay becomes more pliable – a doorway into memory reconsolidation in which emotional tone can be re-tagged while content remains accessible. Default-mode hubs – medial prefrontal and posterior cingulate – participate not as rumination engines but as scene-construction workspaces, allowing safe, vivid simulation of feared or desired futures. In this context, well-timed suggestions function like precision nudges: “feel the urge crest and pass” during smoking imagery, “let the pain be distance rather than danger,” “notice the compulsion without the act.” With repeated trials, cortico-striatal habits adopt the new policy and limbic responses blunt earlier.
1
0

Sep '25 •
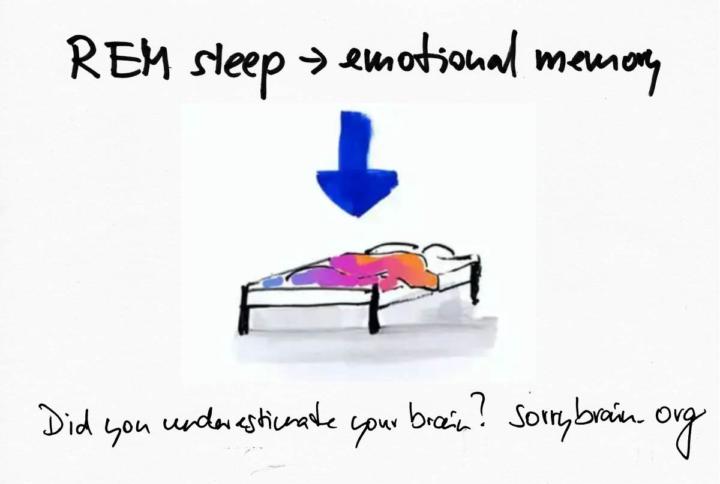
REM sleep and emotional memory consolidation
REM sleep and emotional memory consolidation: “I feel helpless – after hard days I wake in the night, heart racing; arguments replay in technicolor; by morning I’m raw and jumpy.” Here is the bridge to “Now I understand what my brain is able to accomplish.” During REM sleep your brain runs a specialised emotional memory lab – hippocampus, amygdala and medial prefrontal cortex cycle through the day’s charged experiences, extracting what to keep, softening the sting and weaving it into your personal model so you can face tomorrow with steadier affect. Abstract REM sleep and emotional memory consolidation describes a coordinated state – high acetylcholine with low noradrenaline, hippocampal–amygdala–prefrontal communication in theta rhythm, and ponto–geniculo–occipital bursts – that selectively stabilizes and integrates memories tagged with affect. Two complementary outcomes follow: the content of the memory is retained or abstracted, and the affective tone is recalibrated, so tomorrow’s amygdala response is less explosive while prefrontal regulation is better prepared. Sleep before learning, tunes encoding; sleep after learning supports consolidation and integration. When REM is fragmented or scarce, the system under-consolidates context and over-consolidates raw alarm – a profile seen in anxiety, depression and trauma – whereas solid REM predicts better fear extinction, flexible appraisal and social judgment. Evident in Neuroscience and Brain Research Across imaging, electrophysiology and behavioral studies, REM-rich nights bias consolidation toward emotionally salient episodes, increasing next-day recall for the gist while allowing the brain to recontextualize details. Amygdala–hippocampus coupling during REM tracks which items gain a durable foothold; medial prefrontal engagement helps decouple charge from content so recollection the next day is clearer but less incendiary. The REM milieu is distinctive – acetylcholine high, noradrenaline low – which promotes plasticity without the adrenergic “fight-or-flight” stamp. Theta synchrony coordinates hippocampal reactivation with prefrontal evaluation, and PGO waves appear at moments of strong dream imagery when associative linking is maximal. Sleep architecture matters across the night – NREM slow waves stabilize the scaffolding of the memory and down-select noise, while subsequent REM performs affective tuning and network-level integration – so the cycle, not a single stage, carries the benefit. Behaviorally, a consolidated REM period before exposure therapy or difficult social encounters predicts faster fear extinction, fewer intrusive recollections and steadier reappraisal, and REM-selective disruption leaves emotional reactivity artificially high even when neutral memory is intact. Dream mentation is not random nonsense – it is an associative workspace where social and threat models are recombined under low noradrenergic pressure, which is why creative problem-solving and perspective shifts cluster after nights with healthy REM.
0
0
Sep '25 •
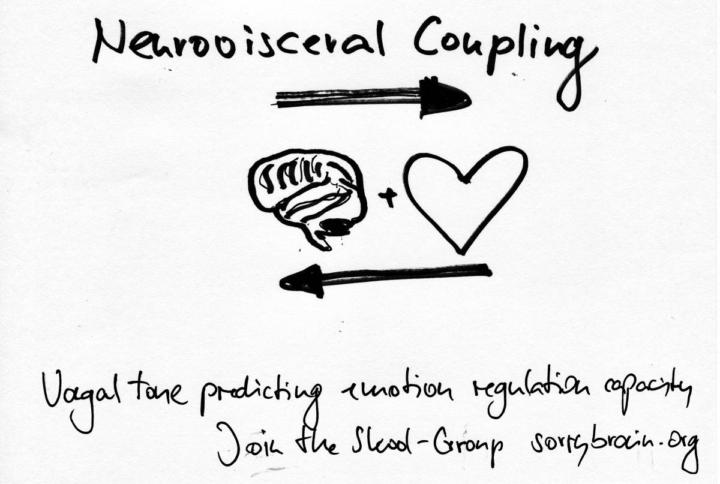
Neurovisceral coupling: vagal tone predicting emotion regulation capacity
“I feel helpless – my heart sprints during conflict, my chest tightens, my stomach flips, I snap or shut down, and I can’t find the brakes.” If that sounds familiar, here is the bridge to “Now I understand what my brain is able to accomplish.” Your body and brain co-regulate through neurovisceral coupling – the dialogue between prefrontal control networks and the vagus nerve that slows the heart, steadies breath and widens choice. The stronger and more flexible that coupling is, the faster you can downshift from surge to steady and the easier it is to steer emotion rather than be steered by it. Abstract Neurovisceral coupling describes how cortical regulators – medial and dorsolateral prefrontal cortex, anterior cingulate and anterior insula – coordinate with the central autonomic network to shape cardiac and respiratory patterns via the vagus nerve. Vagal tone, indexed at rest by high-frequency heart-rate variability and respiratory sinus arrhythmia, predicts emotion regulation capacity – better prefrontal–amygdala control, quicker fear extinction, steadier attention, and prosocial engagement. In stress, vagal output releases its vagal brake, heart rate rises and variability narrows; effective recovery restores variability and calm. Training that raises vagal flexibility – slow breathing around 5–6 breaths per minute, HRV biofeedback, contemplative practice, regular aerobic movement and adequate sleep – tends to improve regulation, while clinical vagus-nerve stimulation can augment treatment in refractory depression and PTSD. Open questions remain about individual baselines, genotype and hormonal moderators, and which protocols best translate acute HRV gains into durable emotion-regulation skills. Evident in Neuroscience and Brain Research At rest, higher high-frequency HRV co-varies with more accurate prefrontal appraisal and fewer impulsive reactions – a signature of a well-tuned parasympathetic system. During affective challenges, people with higher baseline HRV show stronger functional coupling between medial PFC and amygdala, faster return to baseline after stress, and better executive control during distraction. The anterior insula and dorsal ACC act as salience hubs – detecting bodily change and recruiting prefrontal control – and their coherence with cardiac rhythms reflects the tight body–brain loop. Breath-paced HRV biofeedback reliably increases vagal tone in weeks, with parallel reductions in anxiety and rumination and improvements in sleep onset; slow breathing also boosts baroreflex sensitivity, stabilising blood-pressure–heart-rate oscillations that calm arousal. Aerobic exercise and good sleep continuity raise resting HRV and enhance daytime regulation, while sleep loss dampens variability and makes irritability and overreaction more likely. In clinical contexts, vagus-nerve stimulation demonstrates causal leverage – stimulation of afferent vagal fibres alters limbic–prefrontal activity and can lift mood and reduce hyperarousal when other treatments have failed. Together, these findings support a practical rule of thumb: when vagal flexibility is available, the mind has more time to choose.
0
0
Sep '25 •
Books / E-Books Mental Health
Donald Hebb: "The Organization of Behavior: A Neuropsychological Theory 1st edition by Hebb, D.O. (2012) Paperback" (Hebb’s law): https://amzn.to/3Vqny35 Eric Kandel: Principles of Neural Science (memory, synaptic plasticity): https://amzn.to/42avR6W Antonio Damasio: The Feeling of What Happens (somatic markers, emotion-cognition link): https://amzn.to/45IboJ3 Stephen Porges: Polyvagal Theory (nervous system and safety perception): https://amzn.to/45XhG6n Michael Meaney’s work on epigenetics of stress regulation (maternal care changes gene expression): https://www.meaney.lab.mcgill.ca/ https://www.meaney.lab.mcgill.ca/dr-michael-meaney/ MindRooms – https://www.mindrooms.net/ Note: All Links are Affiliate Links.

1-10 of 11

@johannes-faupel-4091
Science-backed blueprints for stubborn success. Strong alternatives to battling doubt, impostor syndrome, and conflicts. Track record.
Active 16h ago
Joined Aug 31, 2025
Frankfurt, Hesse, Germany